Copyright © 2007-2018 Russ Dewey

Animal Information Processing
Comparative psychologists have been very creative in using conditioning techniques to show that animals perform sophisticated information processing. Often the results are similar to human adults or infants.
Is this the return of the "similarity assumption" of Romanes? Not necessarily. There are always two possibilities when non-human animals perform in a way resembling humans:
1. The animal is carrying out the same information processing as humans.
2. The animal is performing the same task in a different way.
Either alternative is interesting to psychologists. The first possibility, that a common mechanism underlies the behavior of different species, might point us toward fundamental or primitive processes widely shared in the animal kingdom.
The second possibility is that different species (or different individuals) perform a task in different ways. That also presents an interesting challenge.
How have comparative psychologists been creative in recent decades? What are the two alternatives, when animals are trained to do "clever" things?
Scientists can compare the different solutions for clues to their advantages and disadvantages. For example, one solution might involve only a few neurons; therefore it might be very efficient. However, the efficient solution may be limited in other ways.
Here are some examples of comparative research showing that different species can perform some of the same tasks as humans, whether or not they do it in the same exact way.
–Kuhl and Miller (1975) showed that a chinchilla was just as good as humans at discriminating complex human speech sounds...an ability once thought to be a unique evolutionary adaptation of humans.
–Brannon and Terrace (1998) showed that chimps could learn to count from 1 to 9 and to identify pictures that contained 1 to 9 different objects.
–Blough (1985) showed that pigeons saw letters of the alphabet much like humans. The pigeons produced a "confusion matrix" (tendency to mistake one letter for another) similar to humans.
–Geiger and Paggio (1975) showed that common houseflies are fooled by the Muller-Lyer illusion (below) perceiving the midpart of the bottom arrow as longer than the midpart of the top arrow.
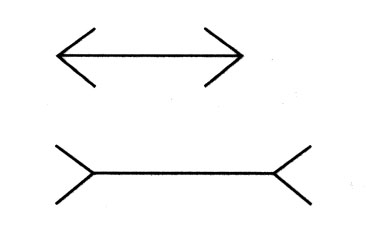
The Muller-Lyer illusion
How did Geiger and Paggio study a housefly's response to the Muller-Lyer illusion?
–Bottle-nosed dolphins have a memory for lists of sounds similar to that of humans (Thompson & Herman, 1977).
–Goldfish can see the "same" color even when spectral composition is radically altered, just like humans (Ingle, 1985).
–Tree swallows follow a logical game-playing or decision-making pattern called Tit-for-Tat (Lombardo, 1985).
–Cats have the same P300 component of the EEG, equated with meaningful interpretation of stimuli, as do humans (Wilder, 1981). So do monkeys (Arthur & Starr, 1984).
–Octopi are capable of observational learning. If allowed to watch another octopus that has been trained to pick one of two targets, the observer will pick the same target (Fiorito and Scotto, 1992).
And the list goes on. Many laboratory tests of human cognitive abilities can be adapted to non-human animals. The result is often the discovery that non-human animals perform similarly to humans.
Specialized Intelligence of Bees
So-called lower organisms often show sophisticated learning abilities related to their unique survival and reproduction needs. Bees and their relatives are especially good at learning the location and appearance of flowers.
What is the waggle dance of the bee?
Bees communicate location information to their fellows with the famous waggle dance first described by Von Frisch (1974). A bee returning to its hive, if it has found flowers, makes a distinctive wiggle of the abdomen and dances up the side of the honeycomb.
The bee's angle from the vertical indicates the angle of the flower away from the sun. A dance 100 degrees to the left of the 12 o'clock position indicates flowers can be found 100 degrees to the left of the sun, as seen from the hive.
If the food is found by flying away from the sun, the bee dances downward rather than upward. The duration of the dance tells other bees the distance of the pollen source (a fast dance indicates food close by). Von Frisch also identified other factors that aid bee navigation, such as odor particles borne by the foraging bee that returns to the hive.
Von Frisch won a Nobel Prize for his work, but many scientists challenged it in various ways. Some suggested that bees responded to sounds rather than the dance, although other researchers believed bees were deaf. Some suggested that bees used odor alone to direct their hive mates, and the waggle dance was irrelevant.
These controversies were put to rest by Kirchner and Towne (1994). They devised robot bees that delivered messages using waggle dances, or by delivering samples of pollen, or by producing a vibrating sound similar to the beating of wings. Among other things, Kirchner and Towne discovered the following:
–Bees can indeed hear, "and their ears are well suited for detecting the sounds associated with the dances."
–"Both sound and dance are needed to communicate information about the location and the food." A bee with clipped wings, which raised the frequency of the sound produced by the dancer, could not recruit other bees.
–Bees in the hive audience may rub their thoraxes against the comb, producing a squeak that vibrates the comb. This causes the dancing bee to stop the dance and dole out samples of food, "so that her audience knows not only the direction and distance to the feeding site but how the food smells and tastes as well."
–A robot bee, dabbed with a faint floral fragrance and "waggled" by a step motor, successfully guides bees to a sugar solution placed in a distant field. (Kirchner and Towne, 1994)
How did researchers test the "waggle dance" theory with robot bees? What other discoveries did they make?
Gould (1986) showed that bees form detailed cognitive "maps." They optimize their routes to flower locations, taking the shortest route when visiting multiple sites.
Bees refuse to respond to a waggle dance that points to the middle of a lake. However, they respond to a waggle dance that points to the opposite shore of a lake.
What is evidence that bees form cognitive maps?
Does it sound unrealistic to suggest that "pictures" and "maps" could be preserved in the tiny heads of insects? The bee brain is tiny, but it is nevertheless a complex system with over 200,000 nerve cells. No wonder they can carry out complex information processing.
---------------------
References:
Arthur, D. L. & Starr, A. (1984). Task-Relevant Late Positive Component on the Auditory Event-Related Potential in Monkeys Resembles P300 in Humans. Science, 223, 186-188.
Blough D. S. (1985) Discrimination of letters and random dot patterns by pigeons and humans. Journal of Experimental Psychology: Animal Behavior Processes, 11, 261-280.
Brannon, E. M., & Terrace, H. S. (1998, October 23). Ordering of the numerosities 1-9 by monkeys. Science, 282, 746-749.
Fiorito, G., & Scotto, P. (1992). Observational learning in octopus vulgaris. Science, 256, 545-547.
Frisch, K. von (1974). Decoding the language of the bee. Science, 185, 663-668.
Geiger, G & Paggio, T (1975). The Muller-Lyer Figure and the fly. Science, 190, 479-480.
Gould, J. L. (1986) The locale map of honey bees: Do insects have cognitive maps? Science, 232, 861-863.
Ingle, D. J. (1985). The Goldfish as a Retinex Animal. Science, 227, 651-654.
Kirchner, W. H. & Towne, W. F. (1994, June). The sensory basis of the honeybee's dance language. Scientific American, pp.74-80.
Kuhl, P. K. & Miller, J. D. (1975). Speech perception by the chinchilla: Voiced-voiceless distinction in alveolar plosive consonants. Science, 190, 69-72.
Lombardo, M. P. (1985). Mutual Restraint in Tree Swallows: A Test of the TIT FOR TAT Model of Reciprocity. Science, 227, 1363-1365.
Thompson, R. K. R. & Herman, L. M. (1977). Memory for lists of sounds by the bottlenosed dolphin: Convergence of memory processes with humans? Science, 195, 501-503.
Wilder, M. B. (1981). Endogenous late positive component on the evoked potential in cats corresponding to P300 in humans. Science, 211, 605-607.
Write to Dr. Dewey at psywww@gmail.com.
Don't see what you need? Psych Web has over 1,000 pages, so it may be elsewhere on the site. Do a site-specific Google search using the box below.